У какого дерева зеленый ствол. Строение древесины лиственных пород
Виды стволов деревьевПрежде чем начать делать какое-либо дерево, необходимо определиться, какой ствол и крона у него будут. Существует несколько видов деревьев, которые можно воссоздать при помощи бисероплетения.
Типы стволов деревьев
Шаккан
Этот стиль подразумевает наклонное дерево. С помощью данного стиля можно сделать любое дерево, которое наклоняется под действие сильного ветра.
Ствол дерева можно сделать прямым, а можно выполнить его немного искривленным.
Буджунги
Данный тип ствола прекрасно подходит для создания стволов деревьев как широколиственных, так и хвойных. Ствол в этом стиле будет выполняться изогнутым, с плавными линиями. Крона у таких деревьев будет располагаться только в самой верхней части деревца, нижних веток нет.
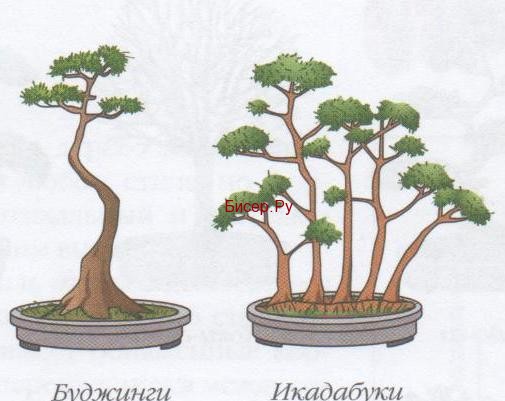
Икадабука
Данный стиль предусматривает выполнение упавших деревьев.
Иси-дзуки
В этом стиле красиво смотрятся клен, сосна, айва. Деревья в этом стиле передают ощущение, что они вросли в камни.

Сокан
Этот стиль прекрасно подходит для всех видов деревьев. Он подразумевает два вида растущих от корней стволов. Один из них будет толще, мощнее другого.
Каабу-дачи
Этот стиль также предполагает два ствола на дереве. Только здесь идет их сдвоение.

Кенгай
В этом стиле лучше всего делать сосны, пираканты, можжевельники. Это не просто дерево, а растущее на очень крутом склоне растение. Оно изгибается не только под собственным весом, но и от камней, которые могли на него попасть при падении вниз с утеса.
Хоки-дачи
Стиль для выполнения деревьев с тонкими ветками (вяз, граб, дзельква).
Как и в хвойных породах, сердцевина лиственных пород образована довольно крупными паренхимными клетками, среди которых иногда встречаются мелкие толстостенные клетки, расположенные поодиночке или небольшими группами и заполненные бурым содержимым; у березы, дуба и ясеня клетки сердцевины могут оставаться живыми до 20-летнего возраста.
Древесина лиственных пород построена более сложно и состоит из большего числа разных элементов, причем на поперечном разрезе радиальное их расположение обнаруживается только у сердцевинных лучей. Сильное развитие отдельных элементов, особенно сосудов, смещает соседние клетки, вследствие чего древесина лиственных пород не имеет того правильного строения, которое характерно для древесины хвойных пород. В состав древесины лиственных пород входят проводящие элементы - сосуды и трахеиды, механические элементы - волокна либриформа и запасающие элементы - паренхимные клетки. Между этими основными видами элементов встречаются переходные (промежуточные) формы; это еще более усложняет строение древесины лиственных пород. На рис. 20 и 21 показаны схемы микроскопического строения древесины дуба (кольцесосудистая порода) и березы (рассеяннососудистая порода).
Сосуды - типичные водопроводящие элементы только лиственных пород представляют собой длинные тонкостенные трубки, образовавшиеся из длинного вертикального ряда коротких клеток, называемых члениками сосудов, путем растворения перегородок между ними. Если при этом в перегородке образуется одно большое округлое отверстие, такая перфорация называется простой. Если после растворения в перегородке остается ряд полос, между которыми расположены щелевидные отверстия, то такая перфорация называется лестничной (рис. 22). У многих пород встречается какой-либо один тип перфораций в сосудах, например: у дуба только простые, а у березы только лестничные. Некоторые породы имеют и те и другие, но и в этом случае преобладает какой-либо один тип перфораций.
Рис. 20. Схема микроскопического строения древесины дуба: 1 - годичный слой; 2 - сосуды; 3 - крупный сосуд ранней зоны; 4 - узкий сосуд поздней зоны; 5 широкий сердцевинный луч; 6 - узкий сердцевинный луч; 7 - либриформ.
После соединения клеток, образующих сосуд, протоплазма и ядро отмирают и сосуды превращаются в мертвые капиллярные трубки, заполненные водой. В крупных сосудах диаметр члеников большой, длина же их нередко меньше диаметра; перегородки между члениками перпендикулярны длине сосуда, перфорации простые. В мелких сосудах диаметр члеников мал, а длина их в несколько раз превышает поперечные размеры; перегородки между члениками сильно наклонены и у многих пород снабжены лестничными перфорациями.
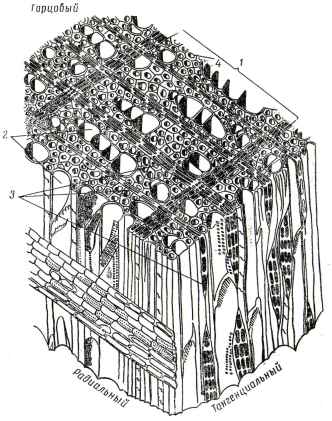
Рис. 21. Схема микроскопического строения древесины березы: 1 - годичный слой; 2- сосуды; 3- сердцевинные лучи; 4 - либриформ.
Таким образом, форма члеников сосудов может быть различной - от веретенообразной в мелких сосудах до цилиндрической или бочкообразной в крупных сосудах; длина их в ранней древесине кольцесосудистых лиственных пород (крупные сосуды) от 0,23 до 0,39 мм, а в поздней древесине (мелкие сосуды) от 0,27 до 0,58 мм. Боковые стенки сосудов разных пород отличаются разнообразием утолщений, возникающих большей частью путем отложения вторичных слоев на первичную оболочку, которая в неутолщенных местах остается целлюлозной и служит для пропускания воды в соседние элементы; утолщенные места обычно древеснеют, так как предназначены для придания прочности стенке сосуда, подверженного давлению со стороны соседних элементов.
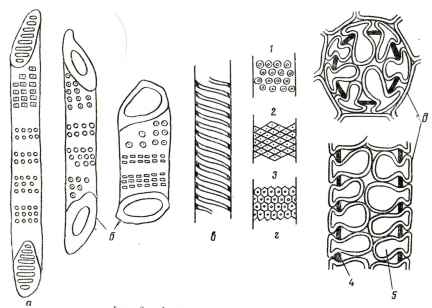
Рис. 22. Детали строения сосудов: а - членик сосуда с лестничной перфорацией; б - два членика сосуда с простой перфорацией; в - спиральный сосуд; г - типы окаймленных пор на стенках сосудов; д - сосуд с тиллами; 1 - округлые поры (береза); 2- ромбовидные поры (клен); 3- многогранные поры (вяз); 4 - стенка сосуда; 5 - тиллы.
Утолщение стенок сосудов делят на кольчатое, спиральное и сетчатое (см. рис. 22). Наименее утолщены кольчатые сосуды. У них утолщения имеют форму колец, расположенных на заметном расстоянии друг от друга; такие сосуды есть только в первичной древесине. Сильнее укреплена стенка сосудов со спиральными утолщениями. У сетчатых сосудов стенка утолщена почти сплошь так, что остаются только поры, заметные в виде частых точек на боковой поверхности сосуда. В древесине большинства лиственных пород встречаются сетчатые, а некоторых пород, например у липы, клена, спиральные сосуды.
В местах соприкосновения стенок с соседним сосудом есть окаймленные поры разной формы, которые отличаются от окаймленных пор хвойных пород меньшей величиной и отсутствием торуса. В местах, где стенка примыкает к паренхимным клеткам, сосуды имеют полуокаймленные поры (окаймление только со стороны сосуда). В местах соприкосновения с клетками сердцевинного луча на стенках сосуда находятся прямоугольные участки, на которых тесно расположены овальные или округлые поры с очень узким окаймлением. В местах соприкосновения с волокнами либриформа стенки сосудов пор не имеют.
Исследования древесины ясеня показали, что сосуды в стволе, отклоняясь от вертикали в тангенциальном и отчасти в радиальном направлениях, сообщаются с соседними сосудами через многочисленные окаймленные поры и перфорационные пластинки. Благодаря указанным конечным и промежуточным контактам в древесине лиственных пород образуется единая пространственно разветвленная водопроводящая система. У некоторых пород с образованием ядра сосуды закупориваются тиллами и выводятся из строя как проводящие элементы. Тиллы представляют собой выросты в большинстве случаев соседних клеток сердцевинных лучей и, редко, древесной паренхимы; они имеют форму пузырей с одревесневшими стенками. Врастание паренхимных клеток в сосуд происходит через поры на его стенках (см. рис. 22).
У некоторых пород тиллы образуются нормально уже после одного или нескольких лет работы сосуда; так, у белой акации и фисташки крупные сосуды частично закупориваются тиллами уже в конце первого года существования. У многих пород тиллами закупориваются обычно сосуды ядра (у дуба, вяза), но и у безъядровых пород в определенных случаях наблюдается сильное тиллообразование (например, в ложном ядре бука). Роль тилл в растущем дереве может быть различной: тиллы закупоривают водопроводящие пути; заполнение сосудов ядра тиллами, особенно толстостенными (у фисташки), повышает твердость древесины; если клетки тилл живые, они играют роль запасающих элементов наряду с древесной паренхимой. В срубленном дереве наличие тилл сильно затрудняет пропитку древесины; например, ложное ядро бука почти не поддается пропитке. Трахеиды у лиственных пород могут быть двух типов: сосудистые и волокнистые (рис. 23). Сосудистые трахеиды - преимущественно водопроводящие элементы, длина которых редко превышает 0,5 мм; по своей форме, размерам, а также по расположению пор они сходны с члениками мелких сосудов; стенки их нередко бывают снабжены спиральными утолщениями. Сосудистую трахеиду можно рассматривать как промежуточный элемент между типичной трахеидой и члеником сосуда.
Волокнистая трахеида в свою очередь представляет собой переходный элемент от трахеиды к волокну либриформа; она имеет форму довольно длинного волокна с заостренными концами, толстую оболочку и малую полость; поры на стенках мелкие, окаймленные, большей частью с отверстием щелевидной формы. От волокон либриформа волокнистые трахеиды отличаются несколько меньшей толщиной стенок, но главным образом наличием ясно окаймленных пор, в то время как у волокон либриформа поры простые. Трахеиды есть в древесине не всех лиственных пород; трахеиды обоих типов есть в древесине дуба, где они приурочены к поздней зоне годичных слоев; волокнистые трахеиды есть в древесине груши и яблони. Либриформ - главная составная часть древесины лиственных пород; у некоторых пород он занимает до 76% общего объема. Волокна либриформа представляют собой прозенхимные клетки веретенообразной формы с толстыми одревесневшими стенками (см. рис. 23), малой полостью и минимальным количеством простых пор на стенках; сбоку поры видны, как узкие щели, расположенные по спирали (косые щелевидные поры). В большинстве случаев заостренные концы волокон либриформа гладкие, но у некоторых пород они расщеплены или имеют зазубрины (у бука, эвкалипта), в результате чего достигается более плотное соединение волокон между собой. Длина волокон либриформа колеблется от 0,3 до 2 мм, а толщина - от 0,02 до 0,05 мм. Рис. 23. Элементы древесины лиственных пород: а - сосудистая трахеида; б - волокнистая трахеида; в - волокно либриформа; г - волокно перегородчатого либриформа; д - тяж древесной паренхимы; е - веретенообразная клетка древесной паренхимы; ж - клетки сердцевинных лучей. Вполне сформировавшиеся волокна либриформа лишены живого содержимого, и полости их заполнены воздухом. Стенки волокон либриформа сильно утолщены в древесине твердых пород (дуба, ясеня, бука, граба и др.) и слабее в древесине мягких пород (липы, тополя, ивы). На рис. 24 показан либриформ с разной толщиной стенок. У некоторых пород, например у кленов, встречаются волокна с менее утолщенными стенками и живым содержимым; эти элементы можно рассматривать как промежуточные между волокнами либриформа и веретенообразными клетками древесной паренхимы. По радиусу ствола размеры волокон либриформа, и толщина, их стенок увеличиваются в направлении от сердцевины к коре, достигают максимума, после чего остаются неизменными или несколько уменьшаются. По высоте ствола длина волокон либриформа и толщина их стенок убывают в направлении от комля к вершине. От количества либриформа и размеров отдельных волокон, главным образом от толщины их стенок, зависят плотность и прочность древесины лиственных пород. Размеры волокон либриформа зависят от условий произрастания: с улучшением этих условий увеличивается длина волокон и толщина их оболочек. Рубки ухода вызывают увеличение количества и длины волокон либриформа. Рис. 24. Фрагменты поперечных разрезов древесины тополя (слева), бука (посредине) и железного дерева (справа): 1 - сосуды; 2- сердцевинный луч; 3, 4 и 5 - волокна либриформа со стенками тонкими, средней толщины и очень толстыми. В древесине некоторых пород (например, тика) встречается так называемый перегородчатый либриформ (см. рис. 23). Его волокна после окончания роста в длину и утолщения оболочек делятся поперечными перегородками на ряд секций; перегородки остаются тонкими и не древеснеют. Таким образом, волокно перегородчатого либриформа несколько напоминает тяж древесной паренхимы, от которого оно отличается характером пор и толщиной боковых (продольных) стенок; кроме того, полости перегородчатого либриформа не имеют содержимого. Сердцевинные лучи. Паренхимные клетки в лиственных породах, как и в древесине хвойных пород, образуют прежде всего сердцевинные лучи, которые у лиственных пород развиты значительно сильнее, нежели в хвойных. Они состоят исключительно из паренхимных клеток, несколько вытянутых по длине луча, с тонкими одревесневшими стенками и многочисленными простыми порами, особенно в тех местах, где клетки луча касаются сосудов или трахеид. По ширине сердцевинные лучи лиственных пород имеют от одного (ясень) до нескольких десятков (широкие лучи дуба, бука) рядов клеток, а по высоте - от нескольких рядов (самшит) до нескольких десятков и даже сотен рядов клеток (дуб, бук). На тангенциальном разрезе однорядные лучи представлены вертикальной цепочкой клеток, а многорядные имеют вид веретена или чечевицы. Строение упоминавшегося выше ложноширокого луча показано на рис. 25. Рис. 25. Сердцевинный луч на радиальном разрезе древесины ивы (слева), и тангенциальный разрез древесины граба (справа): 1 - стоячие клетки; 2 - лежачие клетки; 3 - сосуд; 4 - ложноширокий луч; 5,6 - узкие лучи; 7 - либриформ. У отдельных пород (ивы) краевые клетки, т. е. верхний и нижний ряды по высоте луча, вытянуты поперек луча и называются стоячими (рис. 25); такие лучи получили название лучей разнородных, в отличие от лучей однородных, у которых все клетки по форме одинаковы. Ширина клеток сердцевинных лучей в древесине летнего дуба 15 μ, а высота 17 μ; длина клеток в узких лучах 50-55μ, в широких 69-94 μ. Срединные (по высоте) клетки сердцевинных лучей как у лиственных, так и у хвойных пород по обеим сторонам сопровождаются узкими, заполненными воздухом межклетными ходами, пронизывающими луч по всей длине и через межклетники коровой паренхимы примыкающими к чечевичкам коры; через эти ходы осуществляется газообмен с окружающей дерево атмосферой. Клетки сердцевинных лучей у лиственных пород могут долго оставаться живыми; так, у яблони найдены живые клетки около сердцевины 24-летнего, у бука - 98-летнего, а у граба - даже 107-летнего возраста. Древесная паренхима. Лиственные породы, сбрасывающие листву на зиму, нуждаются в большем, чем хвойные, количестве запасных питательных веществ, необходимых для образования листьев в начале следующего вегетационного периода. Вследствие этого у лиственных пород наряду с большим содержанием (объемом) сердцевинных лучей сильнее развивается древесная паренхима, почти отсутствующая у хвойных пород. Клетки древесной паренхимы собраны в вертикальные ряды и снабжены простыми порами; концевые клетки имеют заостренную форму, благодаря чему весь ряд производит впечатление волокна, разделенного на участки поперечными перегородками (см. рис. 23). Такие ряды паренхимных клеток называют тяжами древесной паренхимы. У некоторых пород (березы, липы, ивы) встречаются веретенообразные паренхимные клетки (веретенообразная паренхима) без поперечных перегородок. Веретенообразная паренхима отличается от трахеиды типом пор и отсутствием спиральных утолщений, от волокон либриформа - толщиной стенок, типом пор и формой окончаний. Древесная паренхима у лиственных пород занимает от 2 до 15% всего объема древесины. У некоторых тропических пород древесная паренхима образует основную массу древесины; такие породы дают особенно легкую древесину (например, бальза). Распределение древесной паренхимы в годичном слое зависит от породы и имеет большое диагностическое значение. Различают следующие основные типы распределения древесной паренхимы: рассеянная (диффузная) паренхима, когда клетки ее распределены по годичному слою более или менее равномерно (береза, бук и др.); приграничная (терминальная) паренхима, когда годичный слой оканчивается одним или несколькими рядами древесной паренхимы (ива, клены и др.); тангенциальная (метатрахеальная) паренхима, когда клетки ее образуют тангенциальные ряды в поздней зоне годичных слоев (дуб, орех грецкий и др.); околососудистая (вазицентрическая) паренхима, когда клетки ее группируются около сосудов. Примерное содержание различных элементов в древесине лиственных пород может быть иллюстрировано данными табл. 6.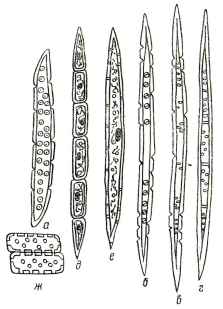
![]()
7.4. Качественные показатели древесных материалов
Древесина хвойных и лиственных пород обладает комплексом свойств, уровень показателей которых обусловлен видом древесной породы, анатомическим строением и составом древесины, степенью оптимизации общей структуры древесного материала, сложившейся в процессе роста дерева в реальных условиях. Особенно значительное влияние на уровень показателей свойств оказывает капиллярное пространство в древесине и проникание воды в капилляры. Одни из них.(первого порядка) бывают относительно крупными, хотя и изменяются с возрастом дерева, - межклеточное пространство, полости клеток, поры в стенках клеток. Другие значительно меньшего диаметра (второго порядка) составляют межфибриллярное пространство и пространство внутри микрофибрилл. Однако и те, и другие, хотя и с разным периодом действия проникшей воды, благоприятствуют набуханию древесных волокон в толщину на 20-30%, в длину - на 1-3% вследствие спиральной структуры волокна .
В древесине всегда содержится некоторое количество влаги. Обычно различают: влагосодержание свежесрубленной древесины - от 35 до 60%, иногда до 100% и более от массы абсолютно сухой древесины; влагосодержание воздушно-сухой древесины - обычно 15-20%, что зависит в основном от относительной влажности окружающего воздуха; влагосодержание мокрой древесины, которое может быть весьма высоким. Например, после длительного пребывания в воде влаги может содержаться до 150% и более от массы абсолютно сухой древесины. Абсолютно сухой древесиной условно считается древесина, высушенная до постоянной массы при температуре 105°С, хотя и в ней всегда содержится небольшое количество влаги.
Влага распределяется в древесине неравномерно: ее больше в комлевой части ствола, в заболонной части, меньше - в ядре, хотя иногда ядровая часть лиственных пород может оказаться влажнее заболони.
Влага бывает свободной, заполняющей внутренние полости клеток, т. е. капиллярное пространство первого порядка, и гигро скопи ческой, находящейся в тончайших капиллярных пространствах второго порядка с возможным переходом в коллоидно-связанное состояние под влиянием взаимодействия ее с веществом дерева. Когда древесина длительное время находится в воздушной среде, насыщенной парами воды, т. е. при относительной влажности воздушной среды, равной 100%, тогда клеточные стенки полностью насыщаются гигроскопической влагой. Этот предел называется точкой насыщения волокон, или пределом гигроскопической влажности. Для древесины различных пород величина предела гигроскопичности колеблется от 23 до 35% массы сухой древесины. В среднем ее принимают равной 30%. К этому времени в волокнах свободной влаги пока еще нет, но набухание достигает своего максимума, причем линейное набухание составляет не выше 6-13%. При набухании наблюдается контракция (сжатие): объем
набухшей древесины меньше суммы объемов древесины до набухания и поглощенной воды. Явление контракции связано со сжатием воды, в частности ее первых порций (примерно до 6%), когда вода имеет повышенную плотность (до 2,6 г/см 3), а объем поглощаемой воды сокращается примерно на 25%. Кроме контракции в процессе поглощения воды возникает также давление набухания (до 100-400 МПа) и выделяется теплота набухания (смачивания). Чем больше в целлюлозе кристаллической части и меньше аморфной, тем меньше набухают волокна и меньше теплота смачивания (набухания), а также сжатие воды.
Явление, обратное набуханию, называется усушкой. Оно выражено уменьшением линейных и объемных размеров древесины при удалении гигроскопической влаги.
Вследствие волокнистого строения древесина имеет разную усушку в различных направлениях. В радиальном направлении она составляет 3-6%, в тангенциальном - в полтора-два раза больше, чем в радиальном, и составляет 6-12%. Усушку вдоль волокон ввиду ее незначительной величины не определяют. Объемную усушку У об вычисляют в процентах без учета продольной усушки по приближенной формуле
где а иb - размеры образца соответственно в тангенциальном и радиальном направлениях;а 0 иb 0 - размеры образца в тех же направлениях в абсолютно сухом состоянии.
Полный объем гигроскопической влаги в древесине (25-30%)
разделяют на адсорбционную, ориентированную водородными связями в наиболее тонких капиллярных пространствах (4-6% от абсолютно сухой древесины),и поглощенную вследствие капиллярной конденсации (20-25% от абсолютно сухой древесины). Остальная свободная влага находится в капиллярном пространстве первого порядка. При высушивании древесины особенно трудно удалить эти 4-6% влаги, так как она прочно связывается водородными связями.
Кроме влажности, гигроскопичности, влагопоглощения, влагоотдачи, характеризующих отношение древесины к водной среде, имеется еще целый ряд физических свойств, обусловливающих качество древесного материала. Для использования древесины в строительных целях наиболее значимыми из них являются: истинная плотность вещества древесины, которая примерно одинакова для разных пород и составляет 1,53-1,55 г/см 3 ;средняя плотность колеблется в широком интервале для различных пород, для одной породы разного возраста или при разном соотношении поздней и ранней древесины. Кроме того, она зависит от влажности и пористости древесины. Как правило, средняя плотность древесины меньше 1 г/см 3 (450-900 кг/м 3), так как объем пор в ней значителен, например у сосны - 50-75, ели - 60-76, дуба - 32-64, липы - 65-75, а объем древесного вещества всего лишь 20-50%.
Так, например, средняя плотность древесины ели составляет 0,37-0,58, сосны 0,3-0,7, дуба 0,51-1,04, березы 0,5-0,75 г/см 3 . Но имеются породы и значительно легче, например, бальзовое дерево (0,1 г/см 3) и значительно тяжелее - железное дерево, бакаут (1,35 г/см 3) и др.
Среднюю плотность древесины с фактической влажностью пересчитывают на стандартную влажность, принимаемую равной 12%:
где ρ o 12 - средняя плотность образца древесины при влажностиW = 12%; k o - коэффициент объемной усушки, который показывает, на сколько процентов изменяется объем образца при изменении его влажности на 1%. У древесины большинства породk o = 0,5 (у березы, бука, лиственницы, грабаk o = 0,6). Его определяют по формуле:k o = V o /(W 1 - W 2 ), гдеV o - объемная усушка;W 1 и W 2 - влажность древесины соответственно начальная и конечная.
Древесина является плохим проводником теплоты, что обусловлено ее пористостью (поры заполнены воздухом). Теплопроводность вдоль волокон значительно больше, чем поперек. Так, например, у сосны вдоль волокон теплопроводность равна 0,35 Вт/(м∙К), а поперек волокон - 0,17 Вт/(м∙К). Она также выше в радиальном направлении, чем в тангенциальном.
Тепловое расширение древесины невелико, причем поперек волокон оно выше в 12-15 раз, чем вдоль (превышая тепловое расширение металлов).
Теплоемкость абсолютно сухой древесины разных пород примерно одинакова: 1,26-1,42 Дж/(г∙К); по мере увлажнения теплоемкость древесины возрастает. Теплота сгорания абсолютно сухой древесины разных пород сравнительно мало различается и составляет 20160-21200 кДж/кг. Понятно, что с увлажнением теплота сгорания древесины сильно снижается.
Температурный коэффициент расширения древесины зависит от породы и направления волокон: вдоль волокон он равен 0,000002-0,00001, поперек волокон - 0,00003-0,00006.
Электропроводность сухой древесины очень мала, особенно при поперечном направлении волокон, поэтому она является хорошим изолятором. Но с увлажнением электропроводность возрастает, что служит основанием для измерения влажности по этому физическому
свойству.
Абсолютно сухая древесина обладает удельным сопротивлением 10 13 -10 15 Ом-м, ее относят к полярным диэлектрикам. По мере увлажнения удельное сопротивление древесины снижается, а при влажности выше предела насыщения клеточных стенок (30%) древесина может обладать ионной проводимостью. Это свойство учитывают, так как диэлектрический нагрев используют в производстве арболита, ДСП и ДВП, при модификации древесины.
Древесина, являясь важным строительным материалом, обладает высокой прочностью при действии сжимающих и растягивающих напряжений, которая находится в прямой зависимости от содержания поздней древесины, пористости и влажности, направления механических сил по отношению к расположению волокон, в чем особенно сильно проявляется ее анизотропия. Анизотропия является следствием медленно развивающейся оптимизации (упорядочения) микро- и макростроения"в условиях роста дерева и максимального сопротивления ствола механическим нагрузкам, с выделением упрочняющих (армирующих) волокон в его тканях. Эти волокна ориентированы по направлениям действия главных напряжений. Вместе с тем они сочетаются с более податливыми волокнами ранней древесины. Можно утверждать, что существенное влияние на анизотропию древесины оказывает ее анатомическое строение (макроструктура), в первую очередь механические ткани. Предел прочности древесины хвойных пород при сжатии в 10, при растяжении в 20-30 раз больше для направления вдоль волокон, а модуль упругости почти в 40 раз больше поперек волокон. Различие "упругих свойств в разных направлениях связано с влиянием сердцевинных лучей, особенно у лиственных пород. Оно проявляется тем больше, чем больше доля сердцевинных лучей как своеобразных лучей жесткости в анатомическом строении древесины.
Ориентированное микро- и макростроение древесины обусловливает ее анизотропию не только при механических, но и при других физических воздействиях. Так, например, коэффициент теплового расширения меньше вдоль волокон и больше - поперек волокон (в тангенциальном направлении). Анизотропия набухания древесины выражена сильнее у хвойных пород, а отношение радиального давления набухания к тангенциальному составляет, по данным Я. Рачковского, для хвойных пород 0,6, а для лиственных - от 0,8 до 1. Вдоль волокон линейное набухание наименьшее (0,1-0,15%), а тангенциальное - наибольшее. Аналогичные анизотропные явления отмечаются, по данным Г.Г. Мудрова, в отношении усушки, теплопроводности, электропроводности и других свойств древесины (рис. 7.12).
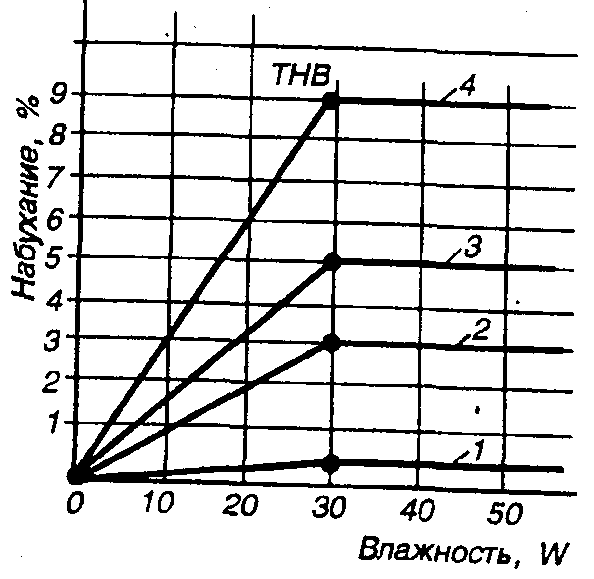
Рис. 7.12. Набухание древесины сосны: 1 - вдоль волокон; 2 - в радиальном направлении; 3 - в тангенциальном направлении; 4 - объемное (ТНВ - точка насыщения волокон)
Предел прочности древесины (с влажностью W в момент испытания) при сжатии вдоль волокон(R сж W ) определяют на стандартных образцах (прямоугольных призмах сечением 20x20 мм и длиной 30 мм) и рассчитывают по формуле
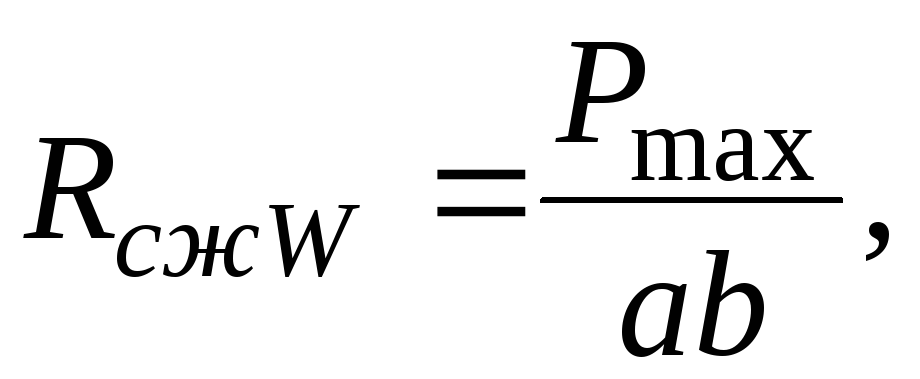 (7.3)
(7.3)
где Р max - максимальная разрушающая нагрузка, Н;a иb - размеры поперечного сечения, м.
Предел прочности при сжатии поперек волокон значительно меньше (10 - 30%) предела прочности при сжатии вдоль волокон.
Наибольшим является предел прочности при растяжении вдоль волокон; он в 2 - 3 раза выше, чем при сжатии вдоль волокон. Предел прочности при статическом изгибе R изг древесины (с влажностьюWn момент испытания) определяют на образцах-балочках размерами 20x20 мм и длиной 300 мм при воздействии двух симметрично расположенных сил и вычисляют по формуле
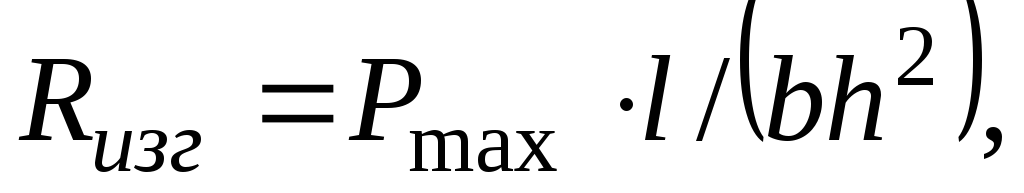 (7.4)
(7.4)
где Р max - разрушающая сила, Н;l - расстояние между опорами (пролет образца-балки), м;b иh - ширина и высота балки, м.
Прочность древесины при скалывании вдоль волокон невелика и составляет примерно 12 - 25% предела прочности при сжатии вдоль волокон.
Прочность древесины значительно понижается с увеличением влажности. Она должна быть приведена к прочности при стандартной влажности 12% по формуле
где R W - предел прочности при влажностиW ,%; W - влажность испытуемой древесины,%;α - поправочный коэффициент на влажность, который показывает, насколько изменяется прочность древесины при изменении влажности на 1% (в пределах влажности от 0 до 30%).
Для сосны коэффициент а при сжатии и изгибе равен 0,04, т. е. материал теряет 4% прочности при увеличении влажности всего на 1%.
Корреляционная связь между прочностью и плотностью древесины, прочностью и процентом поздней древесины выражена в соответствующих эмпирических формулах:
для сосны (7.6)
для дуба (7.7)
где R сж15 - предел прочности при сжатии, кг/см 2 , при влажности 15% (после подсчета пересчитывают на стандартную влажность 12%);ρ o 15 - средняя плотность древесины при влажности 15%, г/см 3 ;m - процент поздней древесины.
Жесткость древесины, ее способность деформироваться под нагрузкой характеризуются модулем упругости: Е = R /ε, гдеR - предел прочности древесины,ε - относительная деформация. Модули упругости при сжатии и растяжении вдоль волокон одинаковы и для сосны составляют 12300 МПа.
Деформативность в направлении вдоль волокон древесины сравнительно низкая. Невелика и твердость ее. Статическую твердость определяют по нагрузке, необходимой для вдавливания в образец древесины половинки металлического шарика радиусом 5,64 мм на глубину радиуса. Тогда площадь отпечатка равна 1 см 2 . Твердость древесины сосны, ели, липы, ольхи составляет 30 - 50 МПа, а более твердых пород - дуба, березы, ясеня, лиственницы и др. - 50 - 100 МПа. Важным технологическим свойством древесины служит способность удерживать гвозди, винты (особенно сосны, ели, ольхи). В лиственных породах (например, дубе) гвозди и винты удерживаются в 16 раз прочнее, чем в древесине хвойных пород. В некоторых породах при забивании гвоздя образуются трещины (бук, дуб, лиственница).
Основные физико-механические свойства древесины хвойных и лиственных пород, применяемых в строительстве, приведены в табл. 7.1.
Таблица 7.1. Основные физико-механические свойства некоторых пород древесины (среднее значение при влажности 12%)
|
Породы дерева |
Средняя плотность, кг/м 3 |
Коэффициент объемной усушки,% |
Предел прочности, МПа, вдоль волокон при |
|||
|
растяжении |
сжатии |
скалывании (радиальном) |
статическом изгибе |
|||
|
Сосна обыкновенная | ||||||
|
Лиственница | ||||||
|
Пихта сибирская | ||||||
|
Лиственные породы |
||||||
В отличие от некоторых других строительных материалов сортность древесных пород устанавливают не по прочности испытуемых образцов, а на основании тщательного осмотра их и оценки имеющихся пороков древесины, нередко значительно снижающих фактическую прочность досок, брусьев, бревен и другой лесопро-дукции. Поэтому целесообразно рассмотреть некоторые, наиболее распространенные, пороки древесины.



